BONUS: Záchrana zelených pokladů: využití metod populační genomiky v ochraně kriticky ohrožených druhů rostlin
Ztráta, degradace nebo fragmentace přirozeného prostředí jsou hlavními faktory odpovědnými za celosvětový úbytek biodiverzity. Areály rostlinných druhů se v čase vyvíjejí a mění. U řady z nich dochází k rozpadu na menší enklávy, což v extrémních případech vede až k úplné izolaci stávajících populací. Dlouhodobá existence takových izolovaných populací je pak závislá především na úrovni jejich genetické diverzity a v neposlední řadě i na nastavení a realizaci vhodné ochrany druhu.
Zmenšováním celkového areálu a snižováním početnosti populací se některé druhy stávají stále vzácnějšími. Jejich přežití můžeme pomoci nastavením vhodných ochranných opatření. Pro to, abychom zjistili, kterým druhům je potřeba věnovat naši zvýšenou pozornost, slouží například tzv. červené seznamy, do kterých jsou dle definovaných kritérií řazeny právě druhy vykazující postupný úbytek přirozených populací. Tyto seznamy nejsou legislativně závazné, a tudíž zařazení některého druhu do národního červeného seznamu automaticky neznamená jeho zákonnou ochranu. Mohou ale sloužit k přípravě nebo aktualizaci právně závazných dokumentů. Všechny planě rostoucí druhy rostlin v České republice jsou obecně chráněny zákonem před potenciálním ohrožením. Druhy, které vyžadují zvláštní ochranu, tzv. zvláště chráněné druhy, jsou zařazeny do vyhlášky. Její součástí je seznam zákonem chráněných druhů. Ty jsou rozděleny do tří kategorií: kriticky ohrožené (C1, např. hořeček mnohotvarý český, šafrán bělokvětý, koukol polní, pelyněk Pančičův, zvonovec liliolistý), silně ohrožené (C2, např. chrpa horská, kosatec nízký, rosnatka okrouhlolistá) a ohrožené (C3, např. hlaváček jarní, dřín obecný, brambořík nachový).
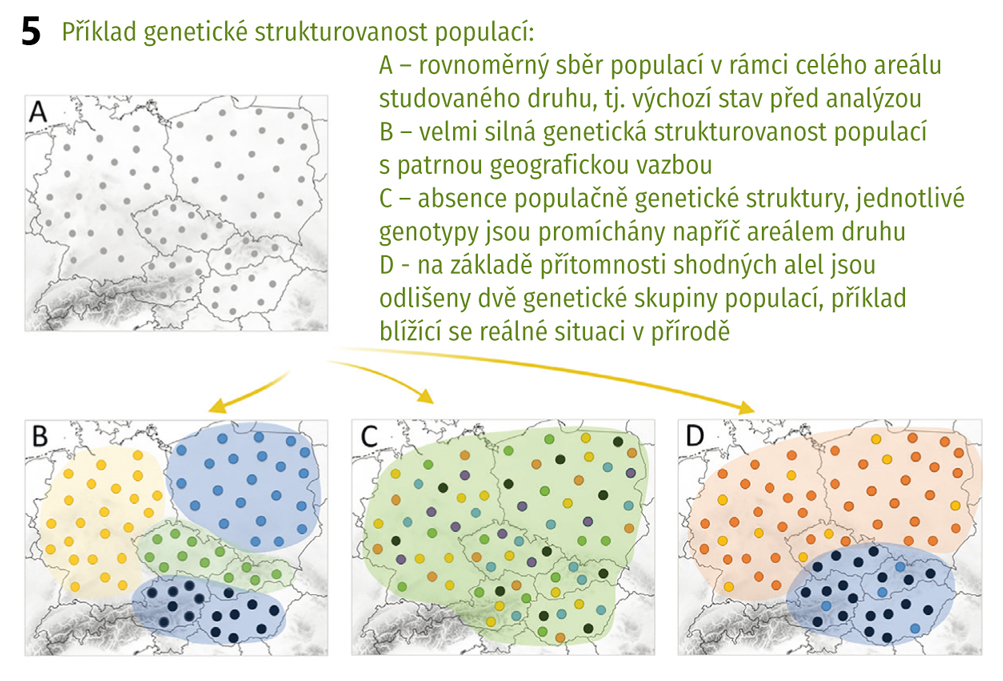
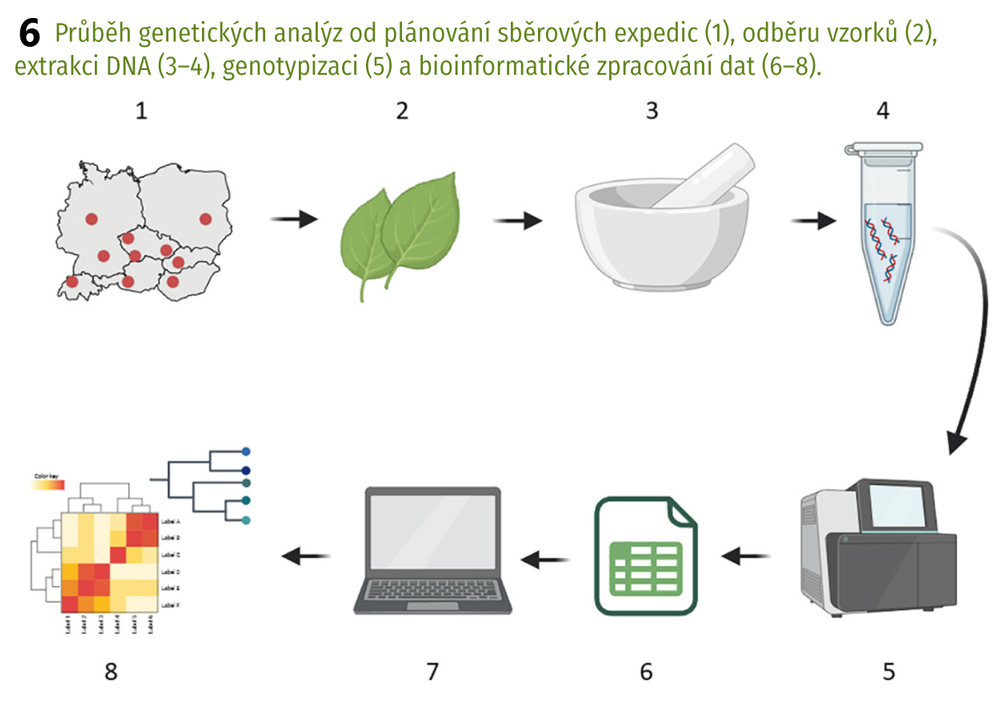
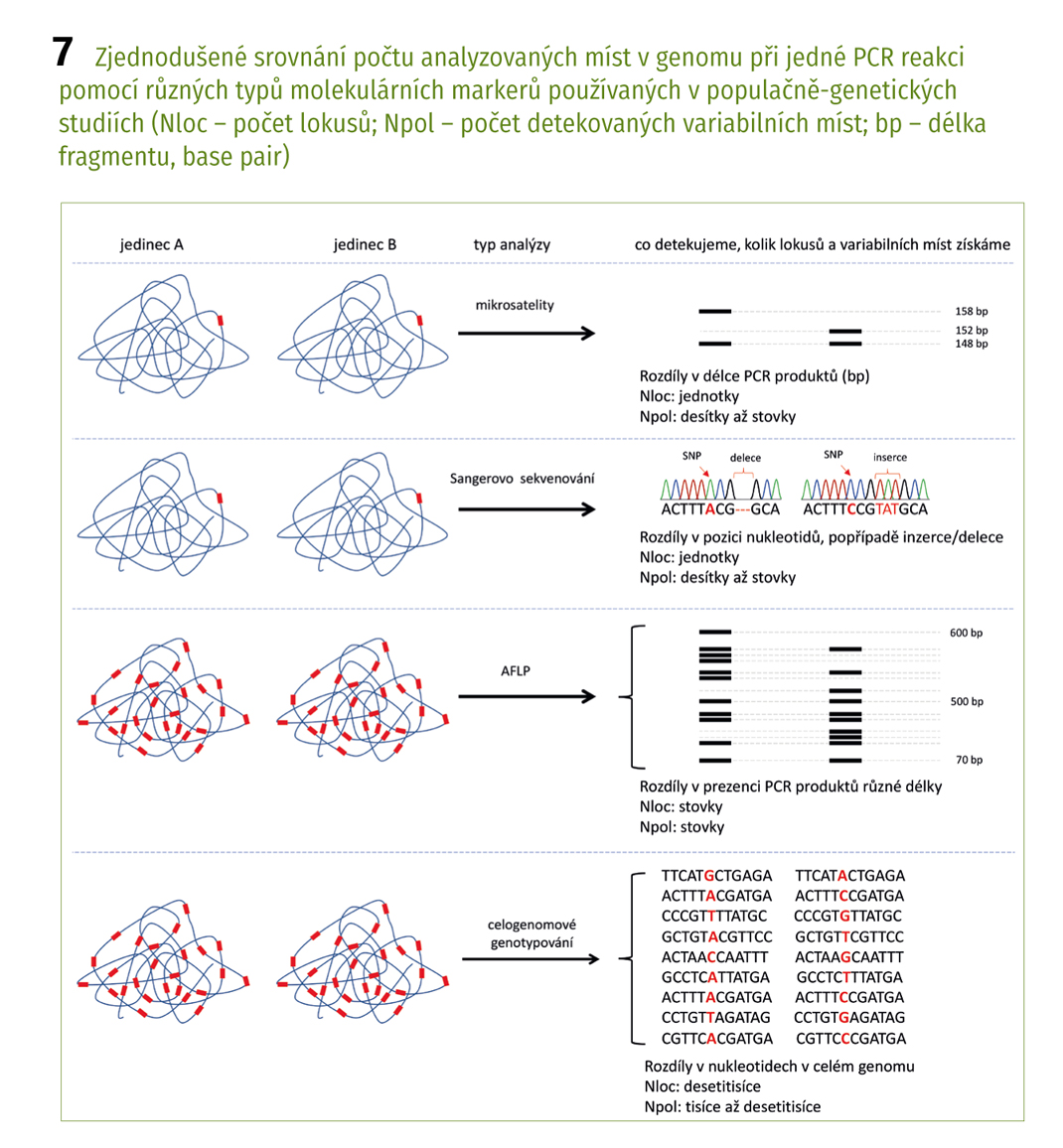
U několika z těchto druhů jsme prováděli populačně genetické studie, jejichž účelem bylo zjistit úroveň genetické variability a vzájemné vztahy ve zbytkových populacích těchto druhů pro ochranářské účely. Jedním z nich byl například pelyněk Pančičův (Artemisia pancicii, C1), panonský endemit rostoucí na několika místech v teplých stepních a lesostepních oblastech jižní Moravy, Rakouska a Srbska. Celosvětově má tento druh do deseti populací, čítajících pouze jednotky až desítky jedinců. Zjistili jsme, že tyto populace jsou geneticky velmi málo variabilní, a ačkoliv například na jižní Moravě od sebe nejsou populace geograficky výrazně vzdálené, nedochází mezi nimi k přenosu pylu (a tím k výměně genetické informace, tzv. toku genů, gene flow), jsou tedy vzájemně izolovány. Postupně došlo v těchto populacích k selekci genotypů (rostlin s unikátní kombinací genů), které jsou nejlépe přizpůsobeny k podmínkám prostředí panujícím na daném místě. Jak tomu ale bude v budoucnu při dnešním trendu stále překotněji se měnících podmínek? Pro druh jako takový by bylo nejspíš nejlepší, kdyby mezi populacemi docházelo alespoň k ojedinělému přenosu genů. Zvýšila by se celková genetická diverzita, tzn. vznikaly by nové genetické varianty, a zároveň by vzrostla šance na přežití druhu v měnících se podmínkách prostředí. V případě, že i nadále zůstanou populace izolovány, může se stát, že v horizontu několika desetiletí postupně vyhynou.
K odlišným závěrům jsme dospěli u zvonovce liliolistého (Adenophora liliifolia, C1; Obr. 1), teplomilného druhu světlých lesů, jehož areál sahá od střední Sibiře až do střední Evropy. V České republice nebyl tento druh snad nikdy hojně rozšířen, ale za posledních 60 let výrazně klesl celkový počet lokalit na pouhých šest, vyskytujících se v severních, středních a východních Čechách. Výsledky našich studií ukázaly, že jedinci v těchto populacích jsou geneticky relativně variabilní a současně jsou si populace vzájemně velmi podobné, a to i přes jejich značnou geografickou izolovanost (na rozdíl od pelyňku). Vyšší celková genetická diverzita tedy dává do budoucna zvonovci větší šanci na přežití a udržení jeho populací bude záviset především na dodržování probíhajícího managementu lokalit (vyřezávání náletových dřevin, ochrana proti okusu vysokou zvěří).