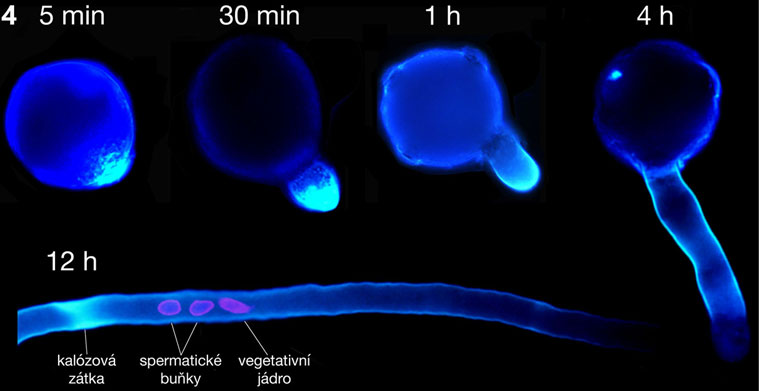
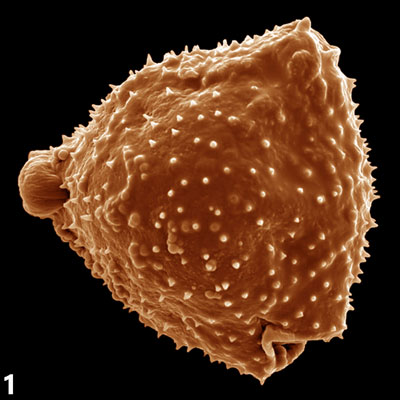
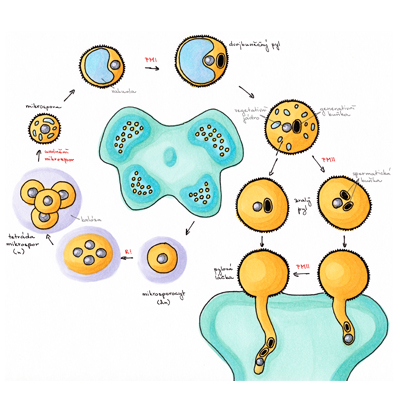
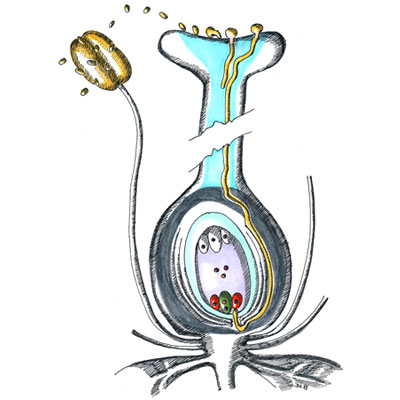
Pro morfologii polarizované pylové láčky je charakteristické asymetrické uspořádání buněčných organel, jež v důsledku proudění cytoplasmy vykazují specifický pohyb popisovaný jako „obrácená fontána“. Růst pylových láček je oscilační a je korelován s oscilací příjmu vápenatých, vodíkových a draselných kationtů a chloridových aniontů. V této souvislosti nepřekvapí objev řady silně exprimovaných a často pylově specifických genů kódujících odpovídající membránové přenašeče, jejichž vyřazení může mít pro správnou funkci pylové láčky fatální důsledky. Apikální (vrcholová) oblast rostoucí pylové láčky nazývaná „clear zone“ obsahuje téměř výhradně transportní vesikuly umožňující sekreci a endofytickou aktivitu potřebnou pro rychlou buněčnou expanzi právě v oblasti špičky spojenou se syntézou buněčné stěny, která je dokončena po reorganizaci jednotlivých komponent v subapikální oblasti. Větší organely bývají uloženy distálněji od subapikálních oblastí cytoplasmy. V této oblasti bývá lokalizován i komplex vegetativního jádra v těsné fyzické vazbě s oběma buňkami spermatickými tak, jak byl vytvořen po ukončení druhé pylové mitosy. Zmíněný komplex se anglicky nazývá „male germ unit“ (MGU) a zastává naprosto zásadní úlohu nejen při dopravě spermatických buněk k vajíčku, ale zejména v přímé komunikaci mezi buňkami a jejich jádry náležejícími k somatické a zárodečné linii a snad i v regulaci jejich genové exprese. Výsledky popsaných ultrastrukturních studií byly potvrzeny i pozorováním živých pylových láček exprimujících bílkoviny asociované s jednotlivými organelami a značené zeleným fluorescenčním proteinem původem z medúz (angl. green fluorescent protein, GFP). Kompartmentace organel je stejně jako jejich pohyb podmíněna zejména propracovanou strukturou aktinového cytoskeletu a jeho zřetelnými strukturálními a dynamickými vlastnostmi v jednotlivých oblastech, jak bylo prokázáno mimo jiné řadou pokusů s látkami inhibujícími polymerizaci aktinových filament. Zatímco úloha mikrofilament v dálkovém vesikulárním transportu i kompartmentaci pylové láčky je poměrně probádaná, funkce mikrotubulů v pylových láčkách zůstává z velké míry nejasnou. V této souvislosti je důležitý fakt, že apikální růst pylových láček krytosemenných rostlin je aplikací inhibitorů tvorby mikrotubulů ovlivněn jen minimálně, avšak přítomnost mikrotubulů je nezbytná pro schopnost udržení směru růstu pylové láčky in vitro. Co se zdá být jasné, je úloha mikrotubulů v udržování komplexu MGU a jeho koordinované migraci v cytosolu pylové láčky. U nahosemenných rostlin se navíc mikrotubuly podílejí na růstu pylových láček koordinovaně s aktinovým cytoskeletem. Mezi typem růstu pylových láček nahosemenných a krytosemenných rostlin vůbec existují podstatné rozdíly, jsou ostatně zcela odlišnými strukturami jak z morfologického, tak z evolučního hlediska.