BONUS: Vývoj pylu: od mikrosporocytu ke zralému pylovému zrnu
Pro životní cyklus rostlin je charakteristické střídání haploidní (gametofytické) a diploidní (sporofytické) generace. Ve vývojové linii cévnatých rostlin je patrná tendence k redukci gametofytu a k jeho stále větší funkční závislosti na sporofytu. U vývojově nejpokročilejších rostlin krytosemenných, jež v současnosti se svými více než 280 000 druhy představují naprostou většinu rostlinných druhů, dosáhla redukce gametofytu svého maxima. Právě ta je spolu s dokonalou ochranou reprodukčních orgánů a využíváním mechanismů výběru životaschopnějšího pylu považována za hlavní příčinu jejich evolučního úspěchu. Většina krytosemenných rostlin vytváří oboupohlavné květy obsahující prašníky i pestíky, kde vznikají a vyvíjejí se samčí i samičí gametofyty. Obě struktury - pylové zrno i zárodečný vak - jsou mikroskopické, sestávající jen z několika málo buněk, a jsou naprosto neschopné vývoje bez podpory okolního sporofytu. Jediným obdobím, kdy se gametofyt semenných rostlin, a to pouze samčí, vyskytuje samostatně, je stadium zralého pylu po opuštění prašníku během svého přenosu na bliznu. Samčí gametofyt představuje zejména ve srovnání se sporofytem jednoduchý a vysoce redukovaný systém, který tak poskytuje unikátní příležitost ke studiu vývojové regulace na mnoha úrovních a také funkčních interakcí mezi různými typy buněk. Právě vývoji samčího gametofytu předurčujícího jej k jeho unikátní roli bude věnován tento příspěvek.
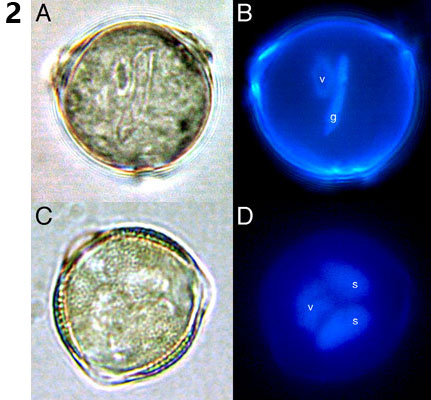
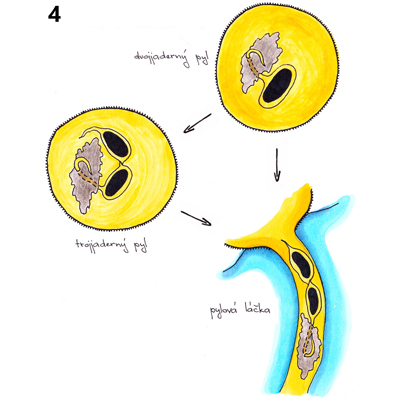
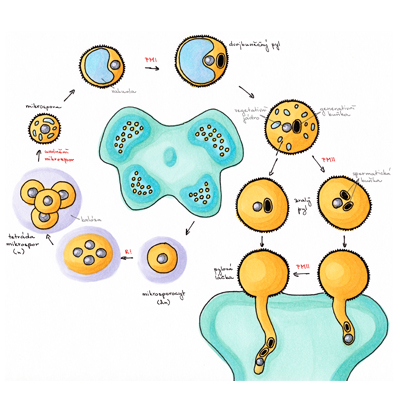
Pylové zrno představuje vysoce redukovanou, dvoj- nebo tříbuněčnou strukturu uzpůsobenou k produkci dvou spermatických buněk a jejich dopravení k samičímu gametofytu, zárodečnému vaku, kde dojde k dvojitému oplození. Diploidní mateřské buňky mikrospor (mikrosporocyty) se nacházejí v prašnících, v prašných pouzdrech obklopených čtyřmi buněčnými vrstvami - tapetem, střední vrstvou, endotheciem a epidermis. Zde mikrosporocyty sekretují unikátní buněčnou stěnu sestávající z ß-1,3-glukanu, kalózy, a poté se během dvou meiotických dělení rozdělí ve čtyři haploidní mikrospory tvořící tetrádu. Mateřské buňky mikrospor navzájem komunikují prostřednictvím cytoplazmatických můstků, což umožňuje synchronizaci jednotlivých meiotických dělení v celém prašníku. II. meiotické dělení je následováno syntézou kalózových stěn také mezi jednotlivými mikrosporami v tetrádě. V době meiotického dělení mikrosporocytů se buňky tapeta diferencují v dvojjaderné polární sekreční buňky postrádající primární buněčnou stěnu, především v místě kontaktu s prašnými pouzdry. Tyto buňky obsahují výjimečné množství ribozomů, mitochondrií, endoplazmatického retikula, Golgiho aparátu a konečně i specializovaných organel bohatých lipidy, tapetozomů, a to zejména v blízkosti plazmatické membrány hraničící s prašnými pouzdry. I buňky tapeta jsou propojeny cytoplazmatickými můstky, což umožňuje koordinaci jejich aktivit.
Recentní studie transkriptomů pletiv prašníků různých rostlinných druhů ukázaly precizní kontrolu aktivity a následné programované buněčné smrti buněk tapeta a koordinaci těchto procesů s vývojem pylu. Mladé mikrospory v tetrádách podstupují bouřlivý vývoj doprovázený syntézou buněčné stěny sestávající z vnitřní intiny a vnější exiny. Po vytvoření částečně zformované exiny jsou mikrospory z tetrád uvolněny. K tomu dochází díky aktivitě směsi enzymů sekretovaných buňkami tapeta, jejíž zásadní složkou je kaláza (ß-1,3- glukanáza) zodpovědná za rozpouštění kalózy. Správné načasování sekrece kalázy je jedním kritických momentů mikrosporogeneze a jeho narušení je jednou z příčin pylové samčí sterility. Volné mikrospory rychle nabývají na velikosti a obsahují větší množství malých vakuol. Tyto časem splývají v jedinou velkou vakuolu vyplňující většinu objemu mikrospory, jejíž přítomnost bývá spojována s přesunem původně centrálně uloženého jádra na přesně určené místo na periferii buňky. Samotný proces přesunu není pasivním jevem jen v důsledku růstu vakuoly, ale procesem dynamickým vyžadujícím aktivní účast mikrotubulů. V tomto období také dochází k dokončení tvorby buněčné stěny syntézou intiny. Buňky tapeta jsou během mikrosporogeneze mimořádně metabolicky aktivní. Sekretují do prostoru prašných pouzder množství bílkovin, lipidů, sacharidů a sekundárních metabolitů, které jsou využívány vyvíjejícími se mikrosporami k syntéze membrán, tvorbě exiny a v neposlední řadě jako zdroj energie. Přes tuto zjevnou důležitost tapeta lze dosáhnout správného dozrání funkčního pylu v podmínkách in vitro již od pozdějších vývojových stadií mikrospor, pochopitelně za přítomnosti základních živin. Mikrospory s jádrem umístěným u stěny podstupují výrazně asymetrické dělení, I. haploidní či pylovou mitózu (PMI). Výsledkem tohoto dělení jsou dvě značně nestejné buňky, buňka vegetativní a generativní, přičemž obě se vyskytují v prostoru ohraničeném původní buněčnou stěnou mikrospory a generativní buňka je posléze zcela pohlcena svou vegetativní sestrou.
Z terminologického hlediska také celý systém v tomto okamžiku přestává být mikrosporou a stává se nezralým pylovým zrnem, nicméně si musíme přiznat, že terminologie je zde natolik komplikovaná a často nejen v různých jazycích různá a neustálená, že její rozbor by vydal na samostatný článek.